1. 最新科研速递

2023年2月28日,白菜网注册领取体验张从刚课题组以及尹航课题组在Cell Reports期刊发表了题为“NF-κB activation enhances STING signaling by altering microtubule-mediated STING trafficking” 的研究论文。该研究发现所有途径(如Toll-like receptor [TLRs]、interleukin-1 receptor [IL-1R]、tumor necrosis factor receptor [TNFR]、growth factor receptors [GF-Rs]、protein kinase C [PKC])激活的NF-κB信号通路,都可以诱导细胞内microtubule解聚,进而阻滞STING通过microtubule迁移到lysosome进行降解的过程,最终高效增强STING介导的先天免疫反应及机体抗病毒能力。这是迄今为止首次在“普适规律”层面,揭示NF-κB能够高效增强STING介导的先天免疫反应,并系统解析了其作用机制,该发现大大拓展了人们对机体免疫体系协同且复杂的工作原理的认识,同时为许多自身免疫性疾病(如STING相关幼年发病性血管病变 [SAVI]、系统性红斑狼疮 [SLE]、肌萎缩侧索硬化 [ALS]、类风湿性关节炎)的潜在发病机制提供了新的见解,并且为自身免疫性疾病、癌症以及感染等众多疾病的治疗提供了重要靶点和全新思路。
2. 研究背景
在1986年,NF-κB复合物体系被诺贝尔奖获得者David Baltimore首次发现1,这开启了先天免疫领域的一个新时代。其后,NF-κB信号通路被证明存在于几乎所有的动物细胞中,并且能够参与细胞对几乎所有外界刺激(如细胞因子、辐射、细菌、病毒、重金属等)的响应,因此在机体炎症反应、自身免疫性疾病、免疫应答、抗肿瘤、抗感染等方面都发挥着重要作用。
20多年后,舒红兵团队2/Glen Barber团队3/蒋争凡团队4以及西南医学中心James Chen(陈志坚)课题组5,6先后发现了STING、cGAS和2’3’-cGAMP。cGAS-STING信号通路的发现是先天免疫领域又一里程碑式的重大突破,破解了“DNA免疫反应”百年谜题。随着研究的深入,科学家发现cGAS-STING通路的重要性已经超出了普通天然免疫通路的范畴,在病原免疫识别、自身免疫反应、抗肿瘤免疫、细胞衰老等诸多方面都具有关键功能。
作为先天免疫系统中至关重要的两条信号通路,NF-κB信号通路和cGAS-STING信号通路之间是否存在互作以及如何互作是一个重要且有趣的科学问题。已经有研究表明STING被激活后会招募TBK1,并磷酸化IRF3,进而诱导IFN表达,同时通过与STING-TBK1-IRF3不同的机制驱动下游NF-κB信号通路的传导7,8。然而,到目前为止,NF-κB信号通路是否影响cGAS-STING信号通路以及如何影响尚不清楚。
3. 研究成果
该研究团队在研究TLR家族的过程中,发现了一个非常有趣的现象:在TLR配体(如HKLM for TLR1/2、LPS for TLR4、R848 for TLR7/8)存在的情况下,STING激动剂(如CDNs、SR-717、DMXAA、HT-DNA、HSV-1)对STING信号通路的激活被显著增强(图1;另见原文Figures 1 and S1)。随后,研究者通过大量的小分子靶向激活/抑制以及基因敲除(Tollip-/-、AKT1-/-、cGAS-/-、MyD88-/-、TBK1-/-、p65-/-、p50-/-)等试验,证明NF-κB是TLRs增强STING介导的先天免疫的核心因子(见原文Figures 2 and S2)。
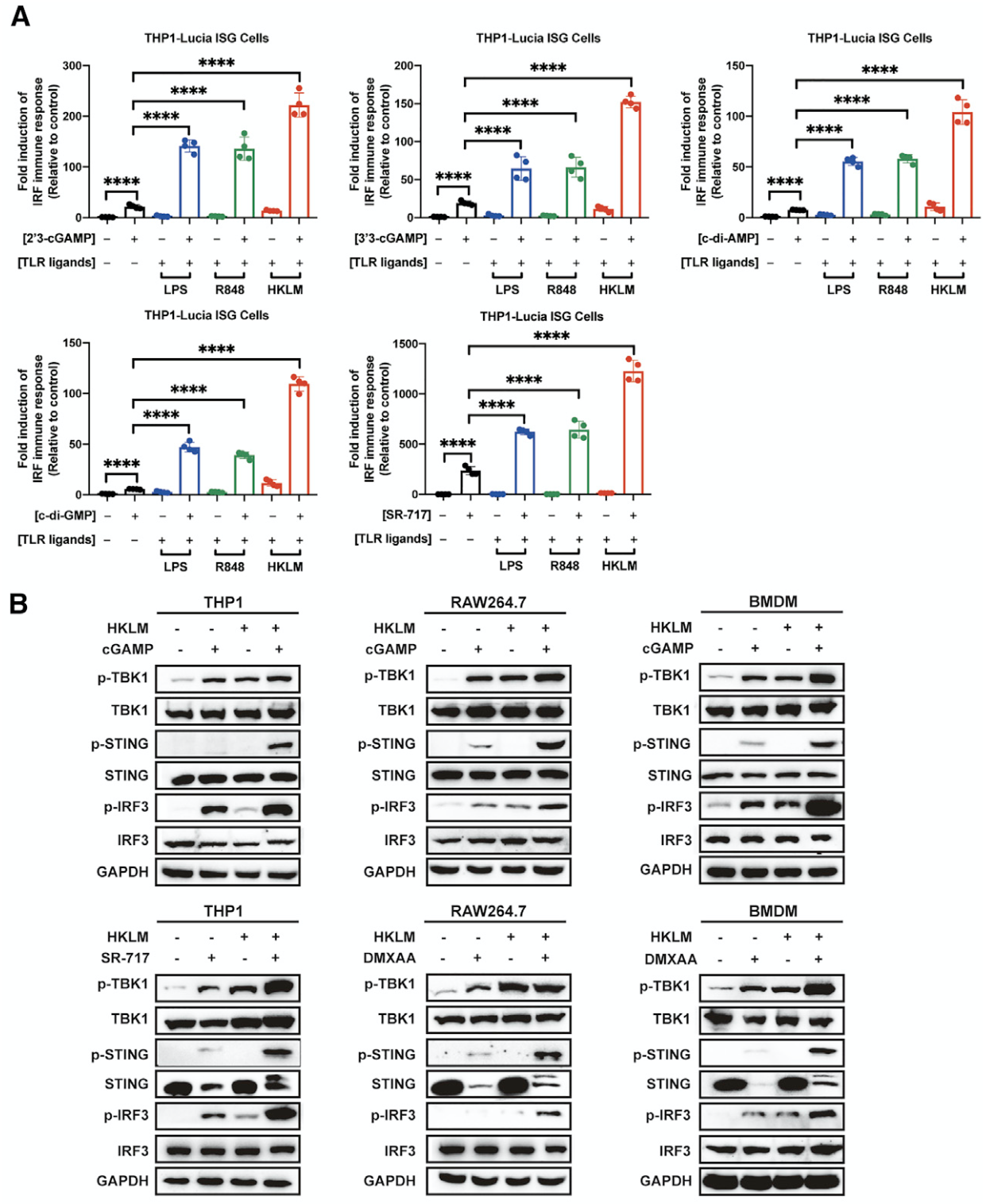
图1. TLR信号通路的激活普遍增强了STING介导的免疫反应
NF-κB信号网络非常庞大和复杂,除了TLRs之外,上游的很多其它受体或蛋白(如IL-1R、TNFR、GF-Rs、PKC)被激活后,也可以通过各自所在的信号轴激活下游的NF-κB。因此,研究者提出一个猜想:如果TLRs可以通过激活NF-κB增强STING介导的先天免疫反应,那么IL-1R、TNFR、GF-Rs、PKC是否也有同样的效果?跟猜想一致,研究者发现所有NF-κB信号通路的激活都能增强STING介导的免疫反应(图2;另见原文Figures 3,S3 and S4)。NF-κB和STING作为先天免疫系统中如此重要的两条信号通路,二者之间存在如此强烈且广泛的互作是惊人的和令人振奋的。
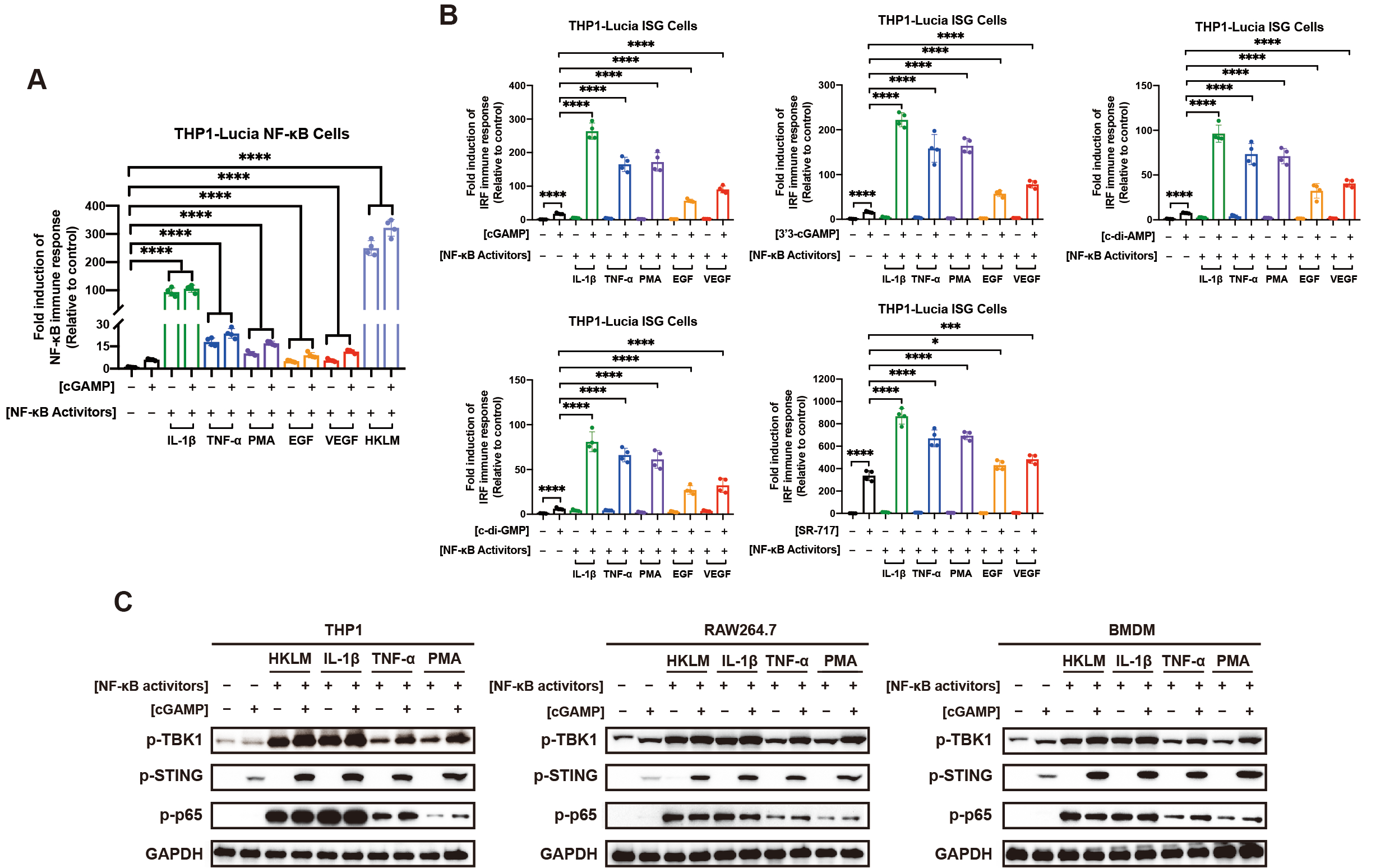
图2. 所有NF-κB信号通路的激活都能增强STING介导的免疫反应
斑马鱼STING的C端尾巴(CTT)含有人类STING并不存在的一段额外延伸序列(CTT-C),这段斑马鱼特有的蛋白序列能够直接招募TRAF6以激活NF-κB 9。这种差异导致STING激动剂对斑马鱼STING的激活强度显著高于人STING。与此一致,通过将鱼CTT-C与人STING蛋白融合,能够大幅提高cGAMP对人STING的激活强度;而剔除鱼STING的CTT-C则迅速降低了cGAMP对鱼STING的激活强度(图3;另见原文Figure S5)。低等脊椎动物(如斑马鱼)缺乏像哺乳动物那样完善的免疫系统,而CTT-C可以通过激活TRAF6-NF-κB信号轴增强STING介导的免疫反应,低等脊椎动物这种独特的STING结构可能有助于保护其免遭一些难以杀死的病原体的侵害。
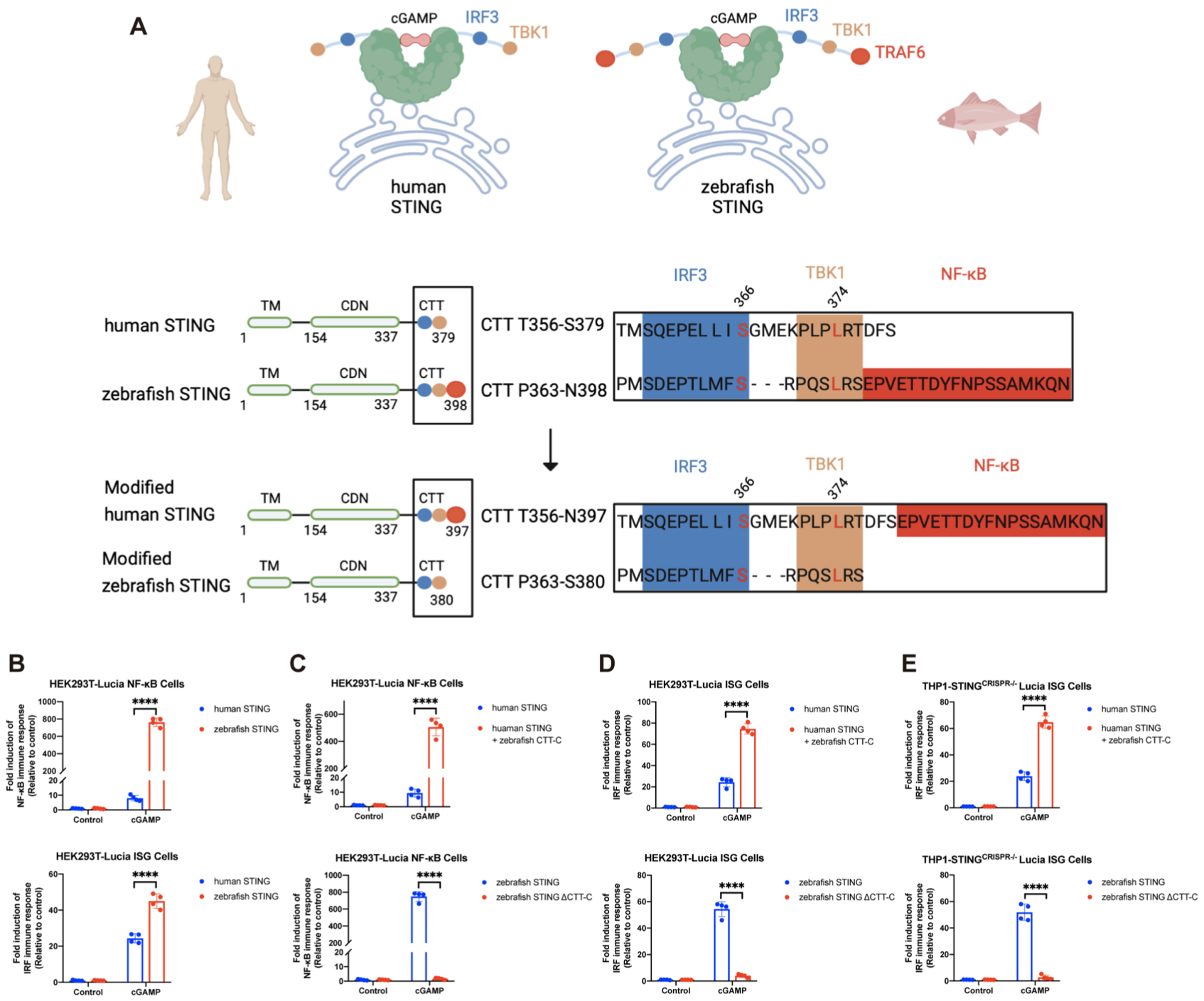
图3. 能够激活NF-κB的斑马鱼STING尾部模块(CTT-C)重新编码人类STING的信号传导
接着,该研究团队详细解析了NF-κB增强STING信号传导的分子机制。通过分析STING蛋白的动态变化,研究者发现NF-κB激活后可以显著抑制STING蛋白的降解,进而导致持续的STING免疫反应。p65的敲除完全消除了IL-1β对STING降解的影响(图4;另见原文Figures 4 and S6)。
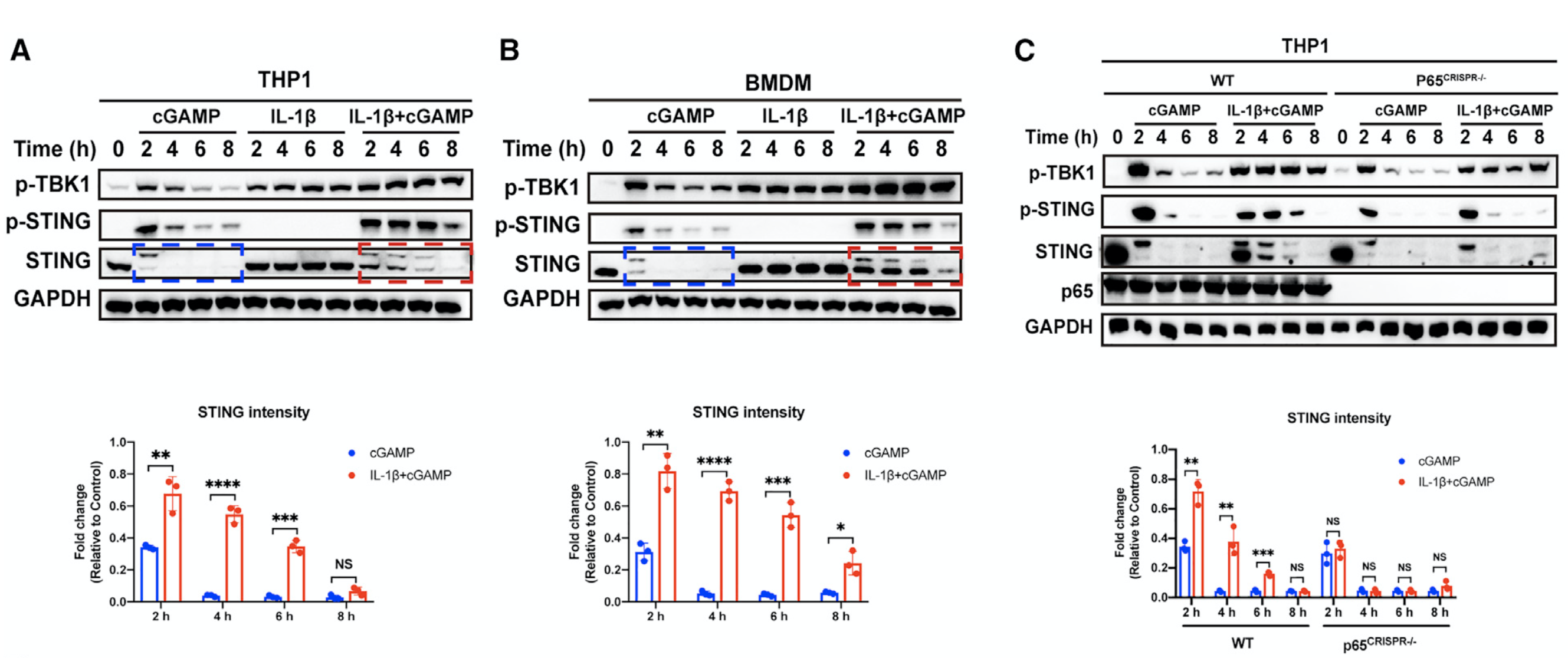
图4. NF-κB的激活阻止了STING的降解,从而延长并增强了STING介导的免疫反应
进一步地,研究者发现STING通过与microtubule结合,沿着microtubule迁移到lysosome进行降解。然而,NF-κB激活会诱导microtubule解聚,进而阻止microtubule介导的STING迁移和降解,从而显著增强了STING介导的免疫反应。敲除p65后消除了NF-κB激动剂对STING迁移和信号传导的影响,却没有消除microtubule抑制剂的影响(图5;另见原文Figures 5 and S7)。综之,NF-κB通过改变microtubule介导的STING trafficking阻止STING降解。这扩展了我们对“机体如何通过microtubule介导的蛋白运输进行精细免疫调节”的理解,并为免疫治疗提供了一个极具吸引力的靶点。
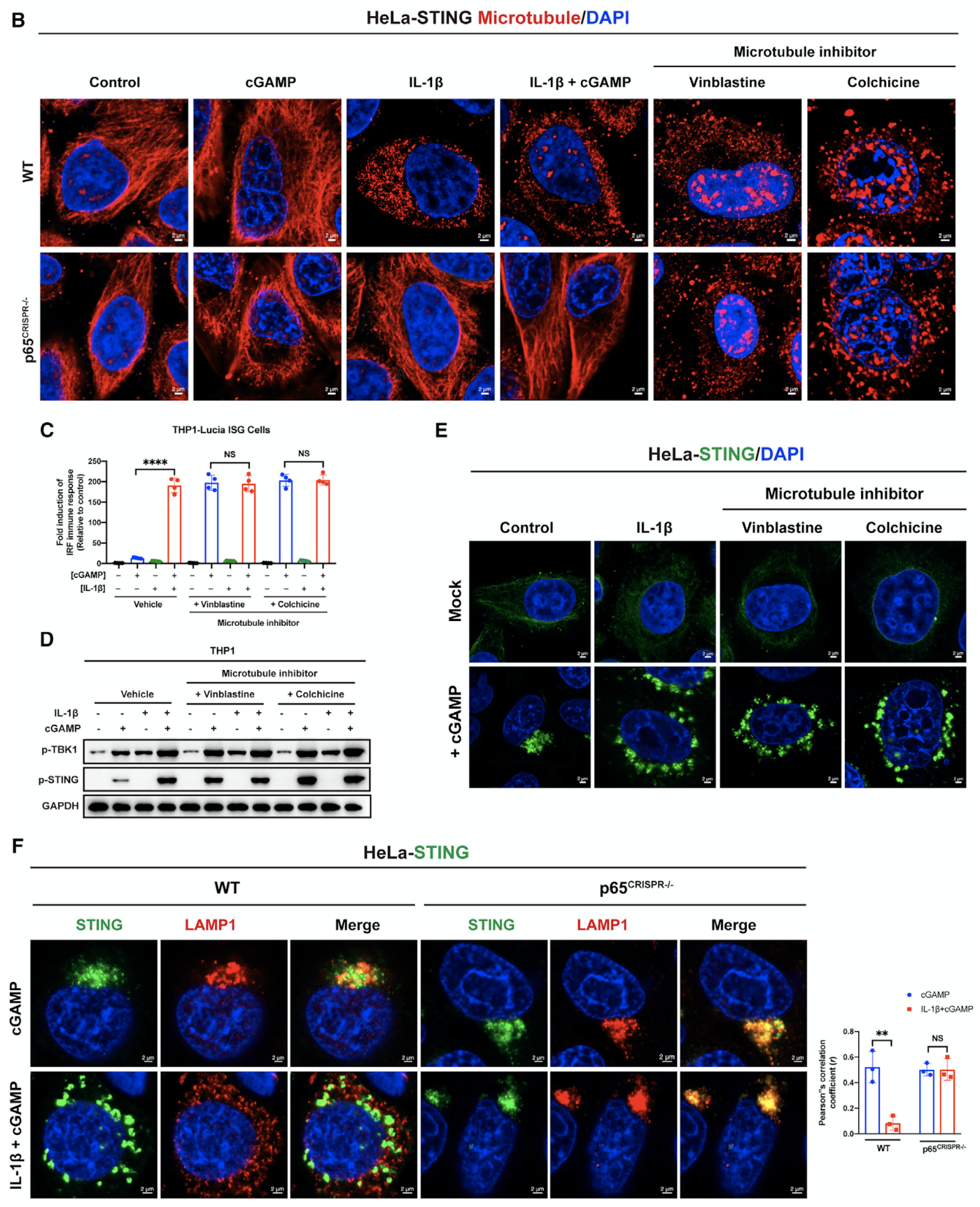
图5. NF-κB通过抑制microtubule介导的STING trafficking来阻止STING降解
SAVI是STING功能性突变导致的一种自身免疫性疾病,但其发病机制目前尚不完全明确。该研究首先发现STING与microtubule结合域在STING胞质区,并且发现在该胞质区与SAVI相关的突变(如F269S、R281E/R284A)会破坏STING与microtubule之间的结合,抑制STING的迁移,进而阻止STING的降解,最终导致STING的持续过度激活(图6;另见原文Figure 6)。这揭示了此类SAVI的潜在发病机制,并为其精准治疗提供了重要启示。
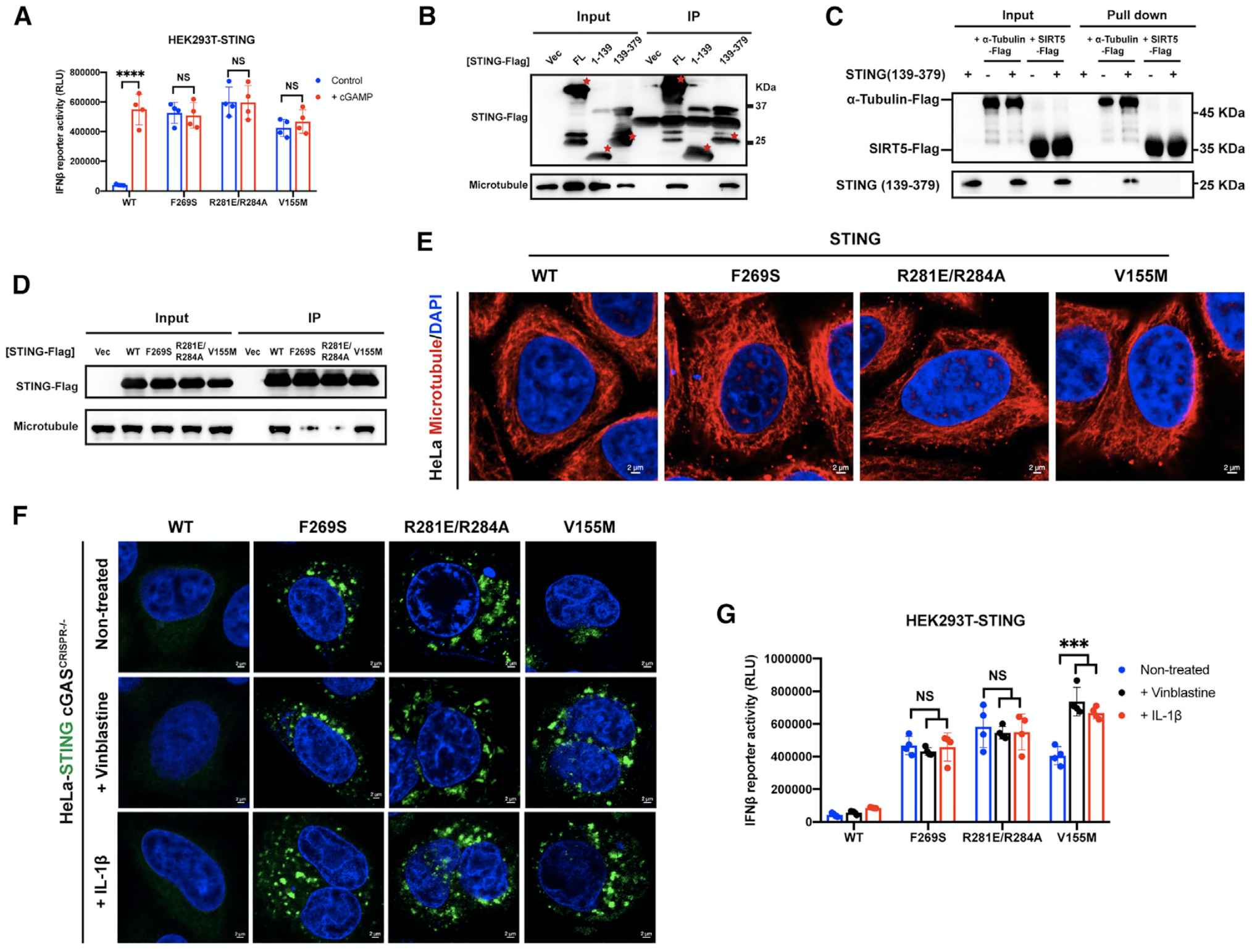
图6. STING的胞质区突变破坏了STING-microtubule结合,并诱发了STING的异常迁移和SAVI
NF-κB激活也显著增强了STING介导的抗病毒免疫。研究者通过体内外试验发现,单独的cGAMP抗病毒效果很有限,但是添加HKLM后却能大幅提高cGAMP诱导的CCL5、CXCL10以及IFNβ的表达,并几乎完全消除了细胞和小鼠脾脏中的病毒感染。然而,在p65-/-的THP1细胞上和MyD88-/-的小鼠(小鼠敲除p65是致死的)上,HKLM对cGAMP介导的免疫反应及抗病毒能力的增强作用完全消失(见原文Figures 7 and S8)。这些数据表明,协同使用NF-κB激动剂和STING激动剂在治疗传染病甚至癌症方面有很大的潜力。
综之,该研究发现NF-κB能够改变microtubule细胞骨架运输网络和细胞器定位,进而改变STING trafficking和降解来增强STING介导的免疫反应。该研究明确了NF-κB信号通路和STING信号通路之间的互作模式和分子机制,扩大了我们对先天免疫调节网络的认识,这对于精准治疗癌症、感染以及自身免疫性疾病具有重要科学价值和指导意义。
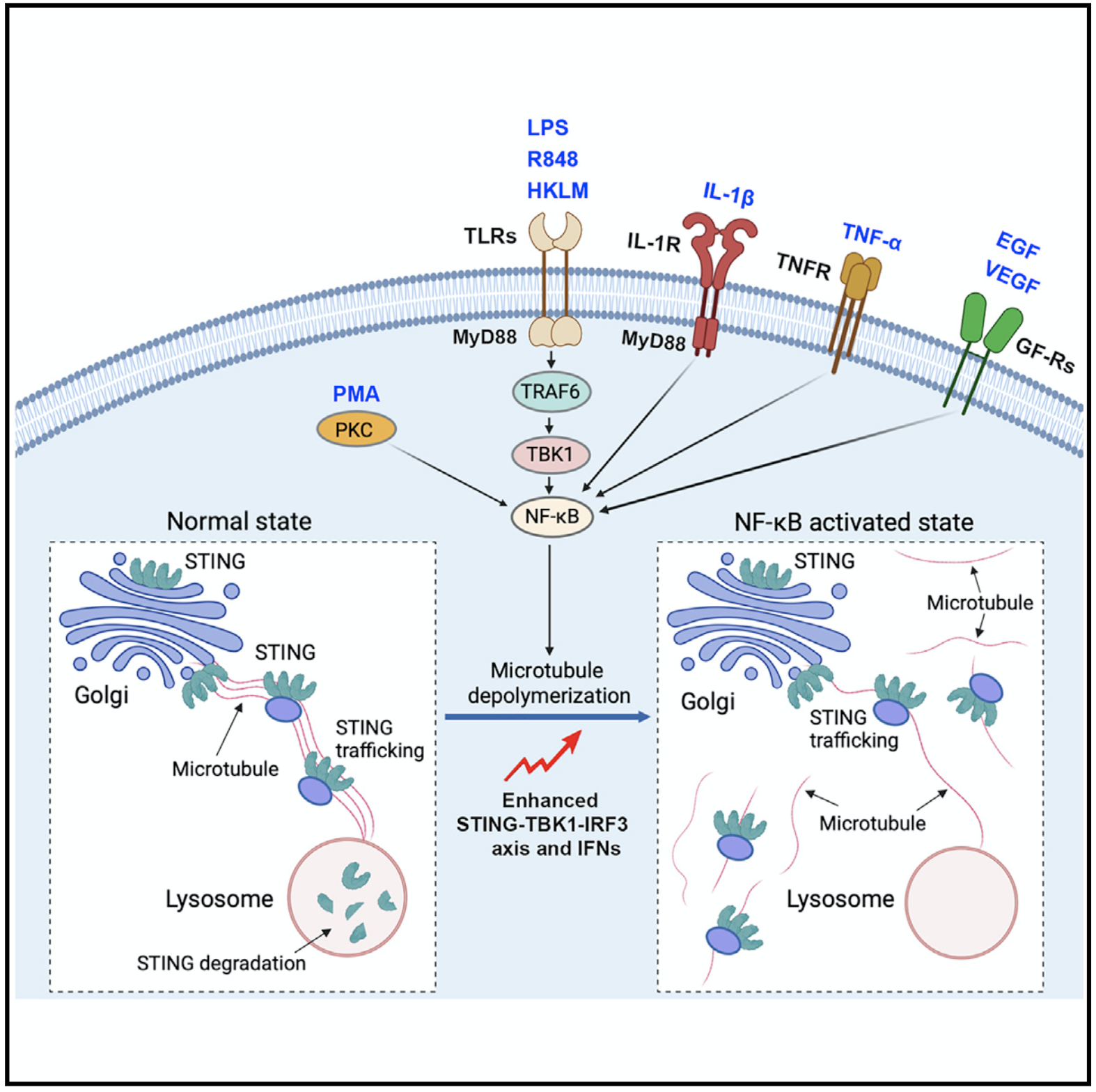
模式图. NF-κB的激活通过改变microtubule介导的STING trafficking来增强STING信号传导
白菜网注册领取体验张从刚研究员和尹航教授为该论文通讯作者,张从刚课题组博士后张璐璐和卫旭彪为该论文第一作者,张从刚课题组王志萌、天津老员工命科学学院刘培源老师以及张从刚课题组多位同学在该课题实施过程中提供了大量帮助,白菜网注册领取体验药学技术中心提供了技术支持。该研究得到了国家自然科学基金项目、清华-北大生命科学联合中心、北京市自然科学基金以及中国博士后科学基金的资助。
参考文献
1 Sen, R. & Baltimore, D. Multiple nuclear factors interact with the immunoglobulin enhancer sequences. Cell 46, 705-716, doi:10.1016/0092-8674(86)90346-6 (1986).
2 Zhong, B. et al. The adaptor protein MITA links virus-sensing receptors to IRF3 transcription factor activation. Immunity 29, 538-550, doi:10.1016/j.immuni.2008.09.003 (2008).
3 Ishikawa, H. & Barber, G. N. STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 455, 674-U674 (2008).
4 Sun, W. X. et al. ERIS, an endoplasmic reticulum IFN stimulator, activates innate immune signaling through dimerization. P Natl Acad Sci USA 106, 8653-8658 (2009).
5 Sun, L., Wu, J., Du, F., Chen, X. & Chen, Z. J. Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 339, 786-791, doi:10.1126/science.1232458 (2013).
6 Wu, J. X. et al. Cyclic GMP-AMP Is an Endogenous Second Messenger in Innate Immune Signaling by Cytosolic DNA. Science 339, 826-830 (2013).
7 Hopfner, K. P. & Hornung, V. Molecular mechanisms and cellular functions of cGAS-STING signalling. Nat Rev Mol Cell Biol 21, 501-521, doi:10.1038/s41580-020-0244-x (2020).
8 Balka, K. R. et al. TBK1 and IKKepsilon Act Redundantly to Mediate STING-Induced NF-kappaB Responses in Myeloid Cells. Cell Rep 31, 107492, doi:10.1016/j.celrep.2020.03.056 (2020).
9 Mann, C. C. D. et al. Modular Architecture of the STING C-Terminal Tail Allows Interferon and NF-kappa B Signaling Adaptation. Cell Rep 27, 1165-+ (2019).

